Catalog Number: LR-039
Manual & Protocol | Material Safety Data Sheets (MSDS)
Lipid rafts are small membrane domains containing a high level of cholesterol and sphingolipids. Lipid rafts are found in the plasma membrane (PM) and internal organellar membranes such as mitochondria-associated membrane (MAMs) and endoplasmic reticulum. They have been implicated in numerous cellular processes including but not limited to signal transduction, membrane trafficking, and protein sorting. Lipid-modified proteins and some transmembrane proteins are concentrated in the rafts while other proteins are excluded. Lipid rafts are also found to be associated with Na+/K+ ATPase on the plasma membrane. Lipid rafts are isolated by sucrose-gradient or OptiPrep gradient traditionally using ultracentrifugation that requires a large amount of starting material. The protocol is tedious and time-consuming. This kit was developed using our proprietary spin-column-based technologies, offering a rapid and easy way to isolate lipid rafts. Total membrane fraction (include PM and organelle membranes) is first isolated and treated with a non-ionic detergent containing buffer, followed by isolation of detergent-resistant fraction by flotation centrifugation using just a tabletop microcentrifuge. Highly enriched total lipid rafts can be obtained from cultured cells/tissues in less than 90 min without using density gradient and ultracentrifugation.
*For isolation of plasma membrane-derived lipid rafts, please refer to MinuteTM Plasma Membrane-Derived Lipid Raft Isolation Kit under Cat #: LR-042
Compare with Other Lipid Raft Isolation kits
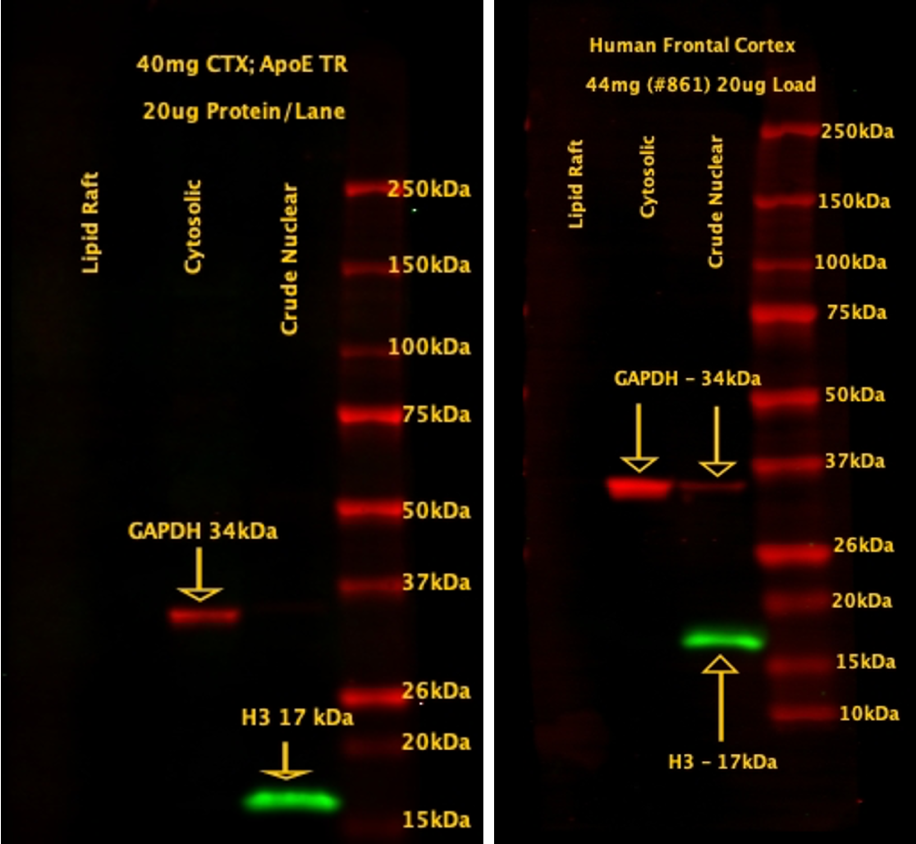
Evaluation of Cross-Contamination of Isolated Lipid Rafts
Lipid rafts were isolated from mouse (TR, left) and human (right) cortex that were probed with H3 and GAPDH. No presence of either in the lipid raft fraction. Human crude nuclear extract has some GAPDH, likely from RBCs. Data: Courtesy of Dr. Max Thorwald, University of Southern California.
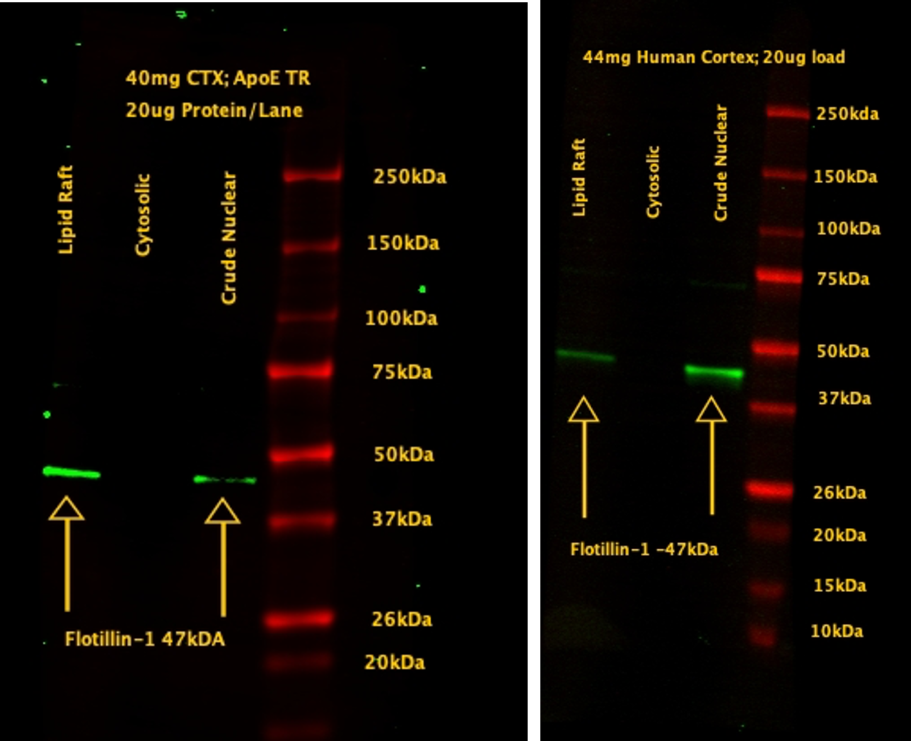
Detection of Lipid Raft Marker in Isolated Lipid Rafts
Lipid raft fractions obtained from isolation kit for mouse (TR, left) and human cortex (right) were probed with Flotillin-1. Lipid raft fractions are positive for Flotillin-1. Data: Courtesy of Dr. Max Thorwald, University of Southern California.
Comparison of Lipid Raft Extractions kit with Ultracentrifugation
Samples were probed with Flotillin-1 on the left. All lipid raft samples probed for H3 and GAPDH samples on the right. The isolation kit yields enriched lipid raft fractions and no visible presence of H3 or GAPDH. Data: Courtesy of Dr. Max Thorwald, University of Southern California.
*TR=transgenic mouse cortex; Human=Human Cortex
Kit includes:
Items
Quantity
Buffer A
15 ml
Buffer B
10 ml
Filter Cartridges
20 units
Collection Tubes (2 ml)
References (11)
1. Wang, M., Liu, J., Tu, Y., Zhao, Z., Qu, J., Chen, K., ... & Wu, C. (2020). RSU-1 interaction with prohibitin-2 links cell-extracellular matrix detachment to down-regulation of extracellular signal-regulated kinase signalling. Journal of Biological Chemistry, jbc-RA120.
2.Thorwald, M. A., Cacciottolo, M., Forman, H. J., Morgan, T. E., & Finch, C. E. (2021). Reductions in ApoE and GPx4 highlight the Alzheimer’s disease lipid raft vulnerability. Alzheimer's & Dementia, 17, e054511.https://doi.org/10.1002/alz.054511
3. Thorwald, M. A., Silva, J., Head, E., & Finch, C. E. (2022). Amyloid futures in the expanding pathology of brain aging and dementia. Alzheimer's & Dementia.
4. Thorwald, M. A., & Finch, C. E. (2022). Alzheimer brains: increased soluble amyloid peptides, but decreased lipid rafts. Alzheimer's & Dementia, 18, e063802.
5. Edwards-Hicks, J., Apostolova, P., Buescher, J. M., Maib, H., Stanczak, M. A., Corrado, M., ... & Pearce, E. L. (2023). Phosphoinositide acyl chain saturation drives CD8+ effector T cell signaling and function. Nature Immunology, 1-15.
6. Thorwald, M., Godoy-Lugo, J. A., Silva, J., Head, E., O'Day, P. A., Morgan, T. E., ... & Finch, C. E. (2023). Alzheimer's Disease associations of ferritin and glutathione with oxidative damage and neuronal loss. bioRxiv, 2023-03.
7. Thorwald, M. A., Godoy‐Lugo, J. A., Forman, H. J., & Finch, C. E. (2023). ApoE4 is associated with higher lipid peroxidation but not protein nitration in AD brains. Alzheimer's & Dementia, 19, e079300.
8. Lei, Y., Yang, H., Xu, X., Tan, Q., Xiang, Y., Tan, T., ... & Xie, P. (2023). Inducing the Localization of HSP70 to Lipid Rafts by Borna Disease Virus 1 Promotes Its Invasion and Host Cells Apoptosis.
9. McGill, C. J., Christensen, A., Qian, W., Thorwald, M. A., Lugo, J. G., Namvari, S., ... & Pike, C. J. (2024). Protection against APOE4-associated aging phenotypes with the longevity-promoting intervention 17α-estradiol in male mice. bioRxiv, 2024-03.
10. Rossi, M., Banskota, N., Shin, C. H., Anerillas, C., Tsitsipatis, D., Yang, J. H., ... & Gorospe, M. (2024). Increased PTCHD4 expression via m6A modification of PTCHD4 mRNA promotes senescent cell survival. Nucleic Acids Research, gkae322.
11. Chaudhary, N., Kasiewicz, L. N., Newby, A. N., Arral, M. L., Yerneni, S. S., Melamed, J. R., ... & Whitehead, K. A. (2024). Amine headgroups in ionizable lipids drive immune responses to lipid nanoparticles by binding to the receptors TLR4 and CD1d. Nature Biomedical Engineering, 1-16.
Minute™ Plasma Membrane/Protein Isolation and Cell Fractionation Kit (50 Preps)
Minute™ Detergent-Free Single Nuclei Isolation Kit (20 Preps)
Minute™ Single Nucleus Isolation Kit for Neuronal Tissues/Cells (20 preps)
Minute™ Total Protein Extraction Kit for Animal Cultured Cells and Tissues (50 Preps)
Minute™ Lysosome Isolation Kit for Mammalian Cells/Tissues (20 Preps)